
Pendant des décennies, le sol sous nos pieds a été traité comme une « boîte noire » dans les modèles climatiques et écosystémiques mondiaux. Nous savions que les microbes du sol étaient responsables de cycles biogéochimiques essentiels, mais leur immense diversité et la complexité de leurs interactions ont conduit à les simplifier en paramètres statiques ou à les ignorer comme « fonctionnellement redondants ». Ma carrière de chercheur a été consacrée à ouvrir cette boîte noire, en utilisant des outils moléculaires émergents et des théories écologiques pour comprendre comment les communautés microbiennes régulent le climat terrestre, répondent aux changements globaux et peuvent même être mobilisées pour la remédiation environnementale.
Ce blog retrace mon parcours, du développement de nouvelles empreintes moléculaires à la cartographie de la biogéographie de microbes spécialisés à l’échelle nationale.
1. Briser la barrière de vitesse : l’ère du T-RFLP Multiplex (2005–2006)
Ma vie de chercheur a débuté par un défi méthodologique fondamental. Au début des années 2000, étudier la structure des communautés microbiennes imposait de choisir entre Bactéries, Champignons ou Archées. Analyser plusieurs groupes simultanément était prohibitif en termes de coût et de temps. Lors de mon Master, ma question de recherche principale était : peut-on développer un test fiable permettant d’étudier plusieurs groupes microbiens simultanément ?
La réponse résidait dans la combinaison de la PCR multiplex avec la sensibilité du T-RFLP (Terminal-Restriction Fragment Length Polymorphism). Cela a abouti au développement du Multiplex T-RFLP (M-TRFLP), une technique permettant la caractérisation simultanée de jusqu’à quatre cibles microbiennes différentes en une seule réaction.
Combler les lacunes : Auparavant, nous manquions de la résolution nécessaire pour observer comment les différents « règnes » microbiens interagissaient dans la rhizosphère. Mes travaux ont validé que le M-TRFLP était aussi robuste et reproductible que les tests individuels, mais nettement plus rentable. En appliquant cet outil à la rhizosphère d’arbrisseaux en Écosse, j’ai constaté que l’habitat et le type de sol — plutôt que l’espèce végétale spécifique — étaient les principaux déterminants de la composition des communautés microbiennes. Ce résultat remettait en cause la croyance selon laquelle les espèces végétales exercent une dominance sélective sur leurs microbes racinaires.
2. Forêts, conversion des terres et rétroactions : le cycle du méthane (doctorat, 2007–2011)
Le méthane ($CH_4$) est 25 à 34 fois plus puissant que le $CO_2$ comme gaz à effet de serre. Si les zones humides émettent du $CH_4$, les sols forestiers des hautes terres représentent le seul puits biologique terrestre du méthane atmosphérique. Au début de mon doctorat, une lacune majeure existait : nous ne comprenions pas pleinement la réponse mécanistique du cycle du méthane aux changements d’utilisation des terres à grande échelle, tels que le boisement et le reboisement.
Ma thèse de doctorat (2011) et les publications associées se sont concentrées sur deux régions clés : la Nouvelle-Zélande et l’Écosse. Les questions de recherche étaient ambitieuses : combien de temps faut-il à un puits de $CH_4$ pour se reconstituer après une perturbation, et ce processus est-il régulé de manière microbienne ?
Résultats clés :
- Le rôle de l’USCa : En utilisant la PLFA-SIP (Stable Isotope Probing) et le T-RFLP, j’ai identifié que l’Upland Soil Cluster alpha (USCa), un groupe de méthanotrophes à haute affinité, est l’ensemble d’espèces clé responsable de la consommation atmosphérique de $CH_4$ à l’échelle mondiale.
- Délais de récupération : Mes travaux ont montré que lors de la conversion de pâturages en forêts, les taux d’oxydation du $CH_4$ se rétablissent beaucoup plus vite que prévu — en 10 à 50 ans.
- Régulation microbienne : J’ai constaté que les changements dans la structure de la communauté méthanotrophe étaient de solides prédicteurs du flux de $CH_4$. Cela a fourni les premières preuves corrélatives que l’« hypothèse de sélection » (où quelques espèces clés pilotent un processus) s’applique à l’écologie microbienne.
- Impact de l’usage des terres : En Écosse, j’ai démontré que la conversion de tourbières et de prairies en forêt de pins pourrait potentiellement neutraliser les émissions nationales de méthane, à condition de préserver le carbone du sol.
3. Réchauffement climatique et réponse respiratoire (2012–2015)
En élargissant mon champ d’investigation, j’ai commencé à étudier la respiration du sol (Rs) — le processus par lequel les microbes du sol libèrent du $CO_2$ dans l’atmosphère. Le réchauffement climatique était censé augmenter Rs, mais les mécanismes sous-jacents faisaient débat : cette augmentation était-elle due à un changement de la structure des communautés microbiennes, à la disponibilité des substrats, ou à une simple accélération métabolique ?
Résultats dans les paysages écossais : Nos recherches sur différents usages des terres (landes, forêts de bouleaux, prairies et forêts de pins) ont montré que la réponse de Rs aux changements d’usage des terres est d’origine microbienne, mais sa réponse au réchauffement climatique ne l’est pas. La réponse à court terme au réchauffement est principalement due à une activité métabolique accrue et à la disponibilité des substrats (la quantité de « nourriture » comme le glucose ou la lignine accessible), plutôt qu’à un bouleversement global de la communauté microbienne. C’est une information cruciale pour les modélisateurs du climat : il faut tenir compte à la fois de l’épuisement des substrats et des taux métaboliques pour prédire les rétroactions futures du $CO_2$.
4. EucFACE et B4WarmED : aux frontières du changement climatique (2015–2021)
Pour comprendre le changement climatique dans le monde réel, j’ai participé à de grandes expériences de terrain comme EucFACE (Eucalyptus Free-Air $CO_2$ Enrichment) en Australie et B4WarmED aux États-Unis. Ces dispositifs nous ont permis de nous interroger : comment l’élévation du $CO_2$ et la modification de la fréquence des précipitations régulent-ils les flux de gaz à effet de serre dans les forêts matures ?
Découvertes clés :
- L’humidité, facteur dominant : Nous avons constaté que l’humidité du sol est souvent un régulateur plus puissant des flux de $CH_4$ et de $N_2O$ que la température ou les niveaux de $CO_2$. Dans les forêts boréales-tempérées, les émissions de $N_2O$ étaient très sensibles à l’abondance des bactéries oxydant l’ammoniac (AOB), tandis que l’absorption de $CH_4$ était limitée par la diffusivité gazeuse (texture du sol) et l’abondance des méthanotrophes.
- Réduction des puits : Nos données EucFACE suggèrent que l’élévation du $CO_2$ pourrait en réalité réduire le puits de méthane forestier dans les écosystèmes arides, ce qui a des implications importantes pour les prévisions futures des bilans de GES.
5. Cartographier l’invisible : biogéographie à l’échelle nationale (2018)
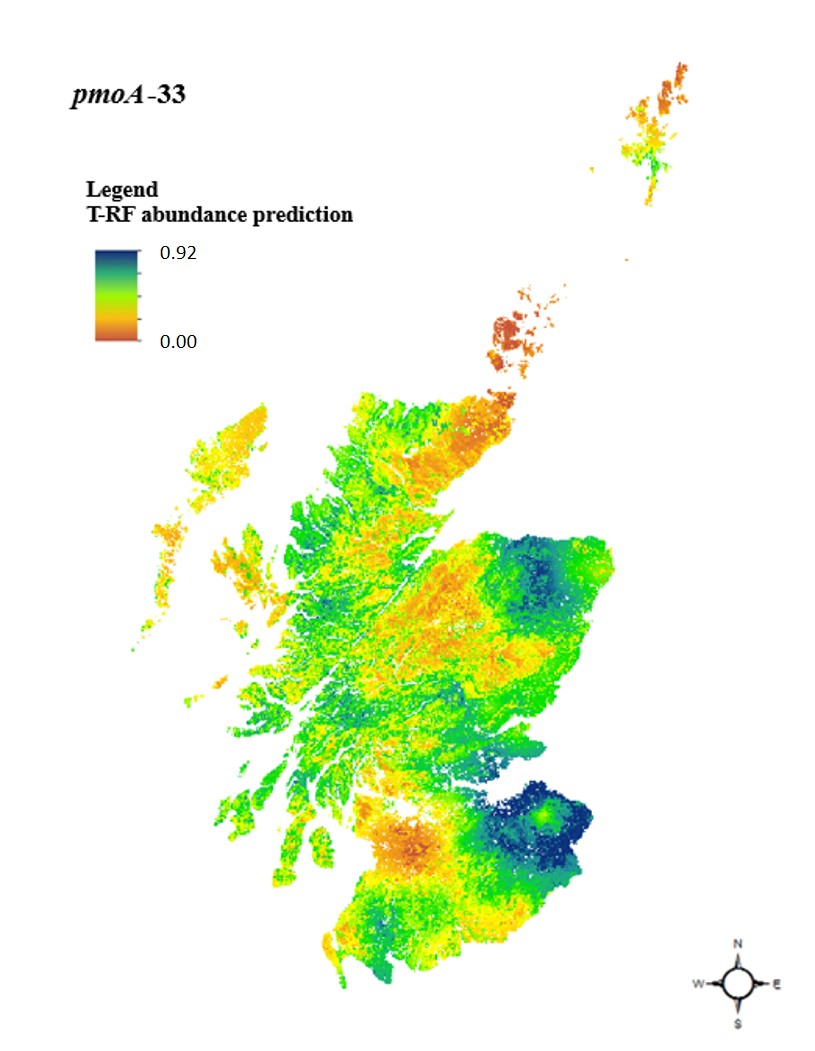
Si nous pouvons cartographier la distribution des plantes et des animaux, pourquoi pas celle des microbes qui pilotent notre climat ? En 2018, j’ai publié un inventaire national des méthanotrophes à travers l’Écosse. Le manque était évident : nous n’avions pas de cartes à grande échelle des groupes fonctionnels microbiens spécialisés.
L’inventaire national écossais : À partir de 697 échantillons de sol et de 62 propriétés climo-édaphiques, j’ai utilisé une modélisation géostatistique hybride pour cartographier les méthanotrophes. Ce travail a révélé un fascinant partitionnement de niche :
- USCa (T-RF 33) dominait dans les environnements plus secs et aérobies, comme les forêts.
- Methylocystis/Methylosinus (T-RF 81) prospérait dans les sols plus humides et riches en matière organique, comme les tourbières.
- Minéraux et métaux : Pour la première fois dans une étude de terrain, j’ai identifié que les ions métalliques du sol (comme Mg, Co et Ni) sont des prédicteurs importants de la distribution de certains méthanotrophes, agissant potentiellement comme des « interrupteurs métalliques » pour l’activité enzymatique.
6. Solutions appliquées : dégradation des pesticides et biochar (2017–2018)
Mes recherches, par le biais d’un de mes étudiants doctorant, ont également porté sur des impacts humains plus immédiats, comme la contamination chimique. Les pesticides organophosphorés comme le Chlorpyrifos (CP) sont largement utilisés mais toxiques. La lacune : nous ne savions pas quelles bactéries du sol spécifiques pouvaient dégrader à la fois le CP et son métabolite toxique, le TCP.
En 2017, mon équipe a isolé de nouvelles souches de Xanthomonas, Pseudomonas et Rhizobium capables de métaboliser efficacement ces toxines comme seule source de carbone et d’azote. C’était le premier rapport de telle polyvalence dans ces genres, offrant un outil puissant pour la bioremédiation des sols agricoles contaminés.
Dans un axe parallèle de recherche appliquée, j’ai étudié la persistance du biochar. Le biochar est souvent présenté comme une « solution miracle » pour la séquestration du carbone. Mes recherches ont précisé que si le biochar peut persister pendant des siècles, son interaction avec la matière organique labile (comme les résidus de cultures) modifie les communautés microbiennes — favorisant spécifiquement les groupes fongiques qui contribuent à stabiliser le carbone à long terme.
Conclusion : de la boîte noire au pouvoir prédictif
En rétrospective, le fil conducteur de mes travaux est la transition de la description à la prédiction. Nous sommes passés d’un simple « les microbes sont là » à l’identification précise des taxa (comme l’USCa) qui pilotent des flux spécifiques ($CH_4$) et de la façon dont les facteurs climo-édaphiques (comme l’humidité et l’usage des terres) déterminent leur distribution.
L’objectif ultime reste : intégrer cette paramétrisation microbienne dans les modèles du Système Terre. En réduisant l’incertitude sur la façon dont les microbes du sol répondent à un monde qui se réchauffe, nous pouvons élaborer de meilleures politiques de gestion des terres et d’atténuation du changement climatique. La « boîte noire » est enfin en train d’être remplacée par une carte complexe, vivante et de plus en plus prévisible du monde microbien.
Avertissement
Ce blog a été entièrement généré par l’usage de l’IA, comme exercice pour tester Google’s NotebookLM: j’ai chargé toutes mes publications comme fichiers