
For decades, the soil beneath our feet was treated as a “black box” in global climate and ecosystem models. We knew that soil microbes were responsible for critical biogeochemical cycles, yet their immense diversity and the complexity of their interactions meant they were often simplified into static parameters or ignored entirely as “functionally redundant”. My career as a researcher has been dedicated to prying open this black box, using emerging molecular tools and ecological theories to understand how microbial communities regulate Earth’s climate, respond to global change, and can even be harnessed for environmental remediation.
This blog post traces my journey from developing novel molecular fingerprints to mapping the biogeography of specialised microbes at a national scale.
1. Breaking the Speed Barrier: The Era of Multiplex T-RFLP (2005–2006)
My research life began with a fundamental methodological challenge. In the early 2000s, studying the structure of microbial communities required choosing between Bacteria, Fungi, or Archaea. Analysing multiple groups was prohibitively expensive and time-consuming. During my MSc, my primary research question was: Can we develop a reliable assay that allows the study of several microbial groups simultaneously?
The answer lay in combining the power of multiplex PCR with the sensitivity of Terminal-Restriction Fragment Length Polymorphism (T-RFLP). This resulted in the development of Multiplex T-RFLP (M-TRFLP), a technique that allowed the simultaneous characterisation of up to four different microbial targets in a single reaction.
Closing the Gaps: Before this, we lacked the resolution to see how different microbial “kingdoms” interacted in the rhizosphere. My work validated that M-TRFLP was as robust and reproducible as individual assays but significantly more cost-effective. Applying this tool to the rhizosphere of dwarf shrubs in Scotland, I found that habitat and soil type — rather than the specific plant species — were the primary drivers of microbial community composition. This challenged earlier beliefs that individual plant species exercised selective dominance over their root microbes.
2. Forests, Land Conversion, and Feedbacks: The Methane Cycle (PhD, 2007–2011)
Methane ($CH_4$) is 25 to 34 times more potent than $CO_2$ as a greenhouse gas. While wetlands emit $CH_4$, upland forest soils represent the only terrestrial biological sink for atmospheric methane. As I transitioned into my PhD, a major gap existed: we didn’t fully understand the mechanistic response of the methane cycle to large-scale land-use changes, such as afforestation and reforestation.
My PhD thesis (2011) and associated publications focused on two key regions: New Zealand and Scotland. The research questions were bold: How long does it take for a soil $CH_4$ sink to recover after disturbance, and is this process microbially regulated?
Key Findings:
- The Power of USCa: Using PLFA-SIP (Stable Isotope Probing) and T-RFLP, I identified that the Upland Soil Cluster alpha (USCa), a group of high-affinity methanotrophs, are the key species responsible for atmospheric $CH_4$ consumption globally.
- Recovery Timelines: My work showed that when pastures are converted to forests, $CH_4$ oxidation rates recover much faster than previously estimated—within 10 to 50 years.
- Microbial Regulation: Crucially, I found that shifts in the methanotrophic community structure were strong predictors of $CH_4$ flux. This provided some of the first correlative evidence that the “Selection Hypothesis” (where a few key species drive a process) applies to microbial ecology.
- Land-Use Impact: In Scotland, I demonstrated that converting bog and grassland into pine forest could potentially neutralise national methane emissions, provided we manage the sites to preserve soil carbon.
3. Climate Warming and the Respiration Response (2012–2015)
As my focus expanded, I began investigating soil respiration (Rs)—the process by which soil microbes release $CO_2$ back into the atmosphere. Global warming was predicted to increase Rs, but the underlying mechanisms were debated: Was the increase due to a change in microbial community structure, substrate availability, or simple metabolic acceleration?
Findings in Scottish Landscapes: Our research across different land uses (moorland, birch woodland, grassland, and pine forest) showed that Rs response to land-use change is microbially explained, but its response to climate warming is not. Instead, the short-term response to warming is primarily driven by increased metabolic activity and substrate availability (how much “food” like glucose or lignin is accessible) rather than a wholesale shift in the microbial community. This was a critical insight for climate modellers: we need to account for both substrate depletion and metabolic rates when predicting future $CO_2$ feedbacks.
4. Navigating the EucFACE and B4WarmED Frontiers (2015–2021)
To understand climate change in the real world, I participated in large-scale field experiments like EucFACE (Eucalyptus Free-Air $CO_2$ Enrichment) in Australia and B4WarmED in the USA. These facilities allowed us to ask: How do elevated $CO_2$ and altered rainfall frequency regulate greenhouse gas fluxes in mature forests?
Key Discoveries:
- The Moisture Overlord: We found that soil moisture is often a more powerful regulator of $CH_4$ and $N_2O$ fluxes than temperature or $CO_2$ levels. In boreal-temperate forests, $N_2O$ emissions were highly sensitive to ammonia-oxidizing bacteria (AOB) abundance, whereas $CH_4$ uptake was limited by gas diffusivity (soil texture) and methanotroph abundance.
- Reduced Sinks: Our EucFACE data suggested that elevated $CO_2$ might actually reduce the forest methane sink in dryland ecosystems, which has significant implications for future GHG budget predictions.
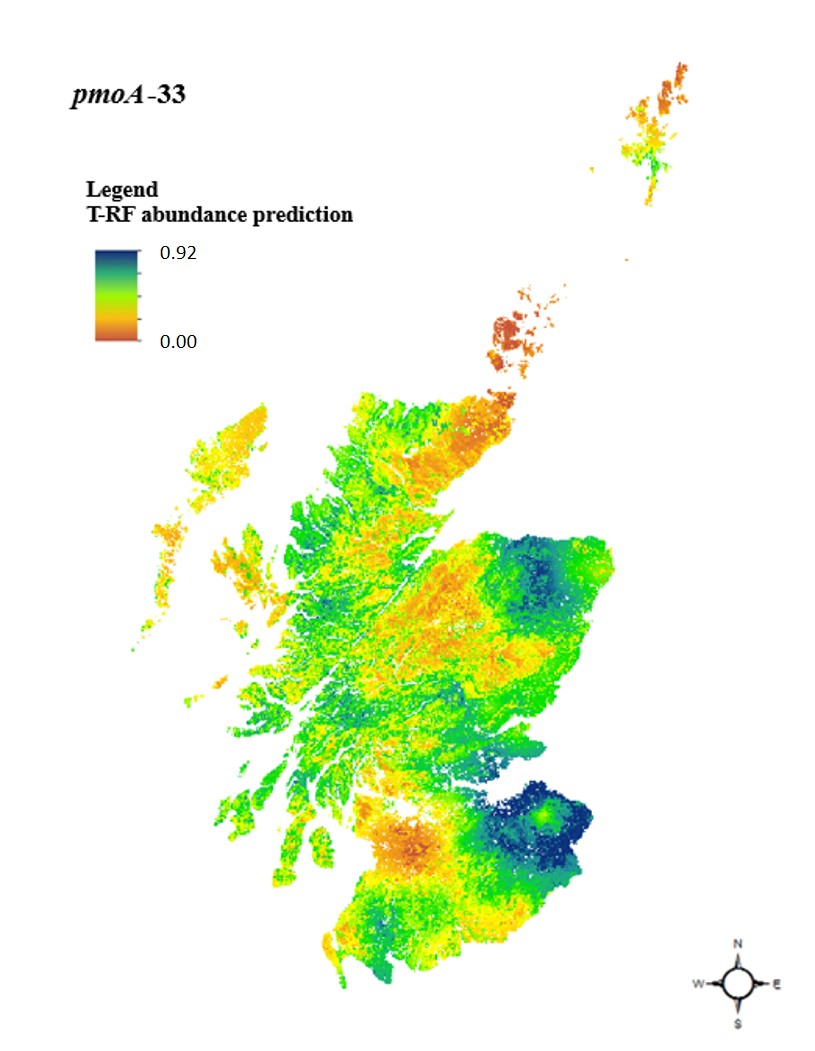
5. Mapping the Invisible: Biogeography at a National Scale (2018)
If we can map the distribution of plants and animals, why not the microbes that drive our climate? In 2018, I published a national survey of methanotrophs across Scotland. The gap was clear: we lacked large-scale maps of specialised microbial functional groups.
The Scottish National Survey: Using 697 soil samples and 62 climo-edaphic properties, I employed hybrid geostatistical modelling to map methanotrophs. This work revealed a fascinating niche partitioning:
- USCa (T-RF 33) dominated in drier, more aerobic environments like woodlands.
- Methylocystis/Methylosinus (T-RF 81) thrived in wetter, organic-rich soils like bogs.
- Minerals and Metals: For the first time in a field study, I identified that soil metal ions (like Mg, Co, and Ni) are important predictors of certain methanotroph distributions, potentially acting as “metal-switches” for enzymatic activity.
6. Applied Solutions: Pesticide Degradation and Biochar (2017–2018)
My research, through the work of one of my PhD students, has also touched on more immediate human impacts, such as chemical contamination. Organophosphorus pesticides like Chlorpyrifos (CP) are widely used but toxic. The gap: We didn’t know which specific soil bacteria could degrade both CP and its toxic metabolite, TCP.
In 2017, my team isolated novel strains of Xanthomonas, Pseudomonas, and Rhizobium that could effectively metabolise these toxins as their sole source of carbon and nitrogen. This was the first report of such versatility in these genera, offering a powerful tool for bioremediation of contaminated farm soils.
In a parallel line of applied work, I investigated biochar persistence. Biochar is often touted as a “silver bullet” for carbon sequestration. My research clarified that while biochar can persist for centuries, its interaction with labile organic matter (like crop residues) shifts microbial communities—specifically favouring fungal groups that help stabilise carbon in the long term.
Conclusion: From Black Box to Predictive Power
Looking back, the recurring theme of my work is the transition from description to prediction. We have moved beyond merely saying “microbes are there” to identifying exactly which taxa (like USCa) drive specific fluxes ($CH_4$) and how climo-edaphic factors (like moisture and land use) determine their distribution.
The ultimate goal remains: incorporating this microbial parameterisation into Earth System Models. By reducing the uncertainty in how soil microbes respond to a warming world, we can develop better policies for land management and climate mitigation. The “black box” is finally being replaced by a complex, vibrant, and increasingly predictable map of the microbial world.
Disclaimer
This blog post was entirely generated by the use of AI, as an exercise to test Google’s NotebookLM: I loaded all my publications as